
In 1957, James Watson and Francis Crick proposed the classical central dogma model of molecular biology: genetic information in biological systems flows from DNA to RNA to protein via transcription and translation, respectively. In 2003, the Human Genome Project successfully sequenced and mapped all 20,000 genes, the DNA instructions for specific proteins, in Homo sapiens. Interestingly, the genes that encode for protein consist of only 2% of the human genome (International Human Genome Sequencing Consortium, 2004). The Encyclopedia of DNA Elements (ENCODE) found that ~70% of DNA is transcribed into non-coding RNA (ncRNA) (Djebali et al., 2012). ncRNAs were long thought to be the “dark matter” or “junk” of the genome since they do not produce proteins like messenger RNAs (mRNAs) and their functions were not well understood (Mattick, 2001).
However, recent research revealed that systemic regulation of genetic expression by ncRNA is integral to biological systems. Aberrant function is linked to a host of diseases, including Alzheimer’s Disease (AD), Parkinson’s Disease, and autism (Nie et al., 2019). Since there are limited treatments for these neurological disorders, ncRNAs and the RNA networks modulating genetic expression are a promising area of research that can expand our arsenal of therapeutic treatments.
However, recent research revealed that systemic regulation of genetic expression by ncRNA is integral to biological systems. Aberrant function is linked to a host of diseases, including Alzheimer’s Disease (AD), Parkinson’s Disease, and autism (Nie et al., 2019). Since there are limited treatments for these neurological disorders, ncRNAs and the RNA networks modulating genetic expression are a promising area of research that can expand our arsenal of therapeutic treatments.
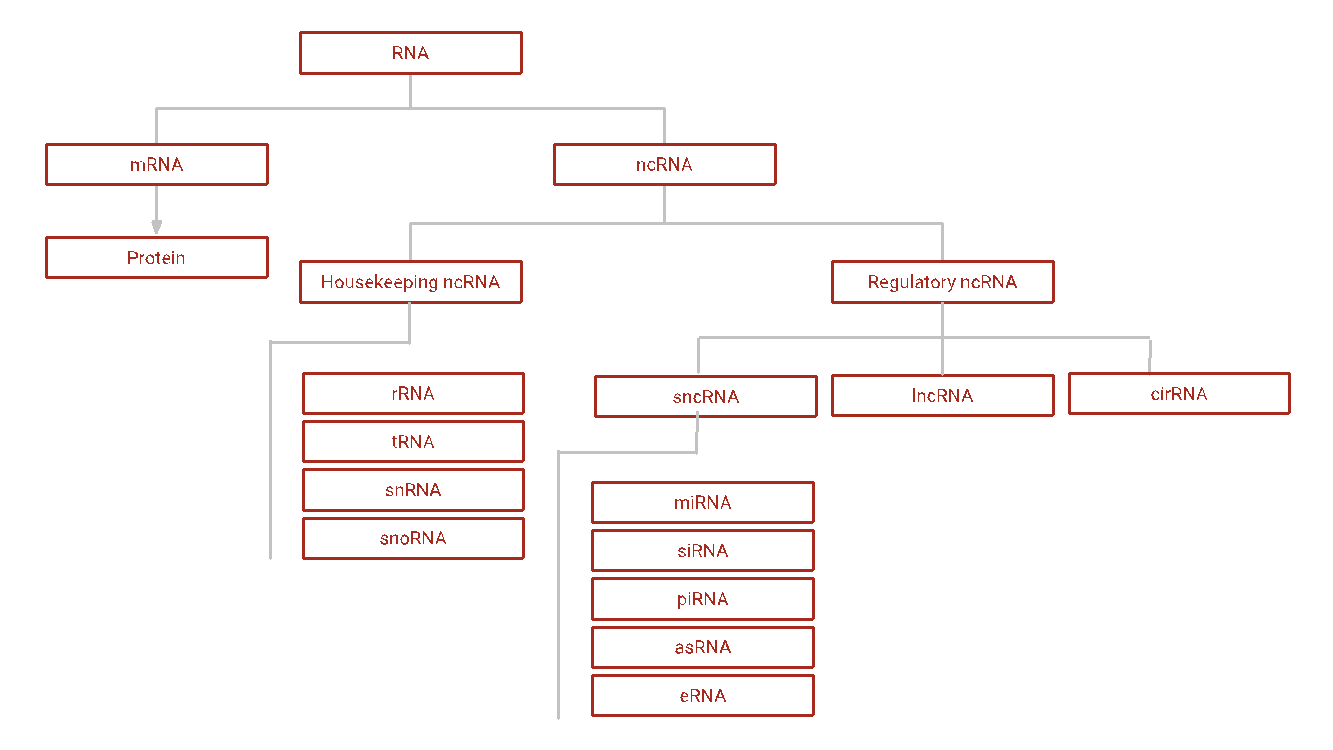
Figure 1: Overview of RNA Classes
|
asRNA = anti-sense RNA
cirRNA = circular RNA eRNA = enhancer RNA lncRNA = long non-coding RNA mRNA = messenger RNA |
miRNA = micro RNA
ncRNA = non-coding RNA piRNA = PIWI-interacting RNA RNA = ribonucleic acid rRNA = ribosomal RNA |
siRNA = small interfering RNA
snRNA = small nuclear RNA sncRNA = small non-coding RNA snoRNA = small nucleolar RNA tRNA = transfer RNA |
There are two types of ncRNAs: housekeeping and regulatory ncRNAs (Baptista et al., 2021). Housekeeping ncRNAs are highly abundant and essential to cellular function. Examples include ribosomal RNAs (rRNAs) that make up the structure of ribosomes, transfer RNAs (tRNAs) that carry specific amino acids to the ribosome during translation, small nuclear RNAs (snRNAs) that catalyze pre-mRNA splicing, and small nucleolar RNAs (snoRNAs) that guide rRNA, tRNA, and snRNA post-transcriptional modifiers.
Regulatory ncRNAs, on the other hand, modulate genetic expression, typically by binding to DNA or RNA to silence expression of these segments of genetic code. Regulatory ncRNAs can be classified by their size. Short ncRNAs (sncRNAs) have less than 200 nucleotides (nt) and are highly diverse. The most studied sncRNAs are micro RNAs (miRNAs), ~20 nt single-stranded RNAs that silence mRNA translation when binding to multiple complementary target sites. miRNAs either induce the degradation of the mRNA or prevent its translation (see figure 2) (Ameres & Zamore, 2013). Imperfect pairing due to their short size allows for less specificity but greater range of inhibitory action. miRNAs serve as biomarkers for various cancers, aging, and neurodegenerative disorders. For example, the neuroprotective miRNA miR-132 has been reported to be severely downregulated in AD and other tauopathies. The overexpression of miR-132 via miR-132 mimic oligonucleotides can rescue tauopathic AD-like rodent neurons from neurodegeneration (Fatimy et al., 2018).
Regulatory ncRNAs, on the other hand, modulate genetic expression, typically by binding to DNA or RNA to silence expression of these segments of genetic code. Regulatory ncRNAs can be classified by their size. Short ncRNAs (sncRNAs) have less than 200 nucleotides (nt) and are highly diverse. The most studied sncRNAs are micro RNAs (miRNAs), ~20 nt single-stranded RNAs that silence mRNA translation when binding to multiple complementary target sites. miRNAs either induce the degradation of the mRNA or prevent its translation (see figure 2) (Ameres & Zamore, 2013). Imperfect pairing due to their short size allows for less specificity but greater range of inhibitory action. miRNAs serve as biomarkers for various cancers, aging, and neurodegenerative disorders. For example, the neuroprotective miRNA miR-132 has been reported to be severely downregulated in AD and other tauopathies. The overexpression of miR-132 via miR-132 mimic oligonucleotides can rescue tauopathic AD-like rodent neurons from neurodegeneration (Fatimy et al., 2018).
Figure 2: miRNA mechanism of genetic silencing
The miRNA (blue) binds to its target site on the mRNA (red) with the help of the RNA-induced silencing complex protein (RISC) (dark blue). The target mRNA can be directly cut by RISC and thus degraded. Alternatively, RISC may block the translation of the target site.
The miRNA (blue) binds to its target site on the mRNA (red) with the help of the RNA-induced silencing complex protein (RISC) (dark blue). The target mRNA can be directly cut by RISC and thus degraded. Alternatively, RISC may block the translation of the target site.
Unlike miRNAs, the ~20 nt double-stranded small interfering RNAs (siRNAs) specifically silence a single target sequence of RNAs via complete complementary binding to mRNA. Another class of sncRNAs called PIWI-interacting RNAs (piRNAs) utilize a protein called PIWI to cleave target RNA. Smaller ncRNAs called antisense RNAs (asRNAs) are single-stranded RNAs ranging from 19-23 nts that typically block translation by binding to a complementary mRNA (Xu et al., 2018). Enhancer RNAs (eRNAs) are a new class of ncRNAs that consist of RNA molecules with ~150 nts, facilitating genetic enhancers to promote transcription (Sartorelli & Lauberth, 2020).
Long ncRNAs (lncRNAs) have more than 200 nt and constitute most non-coding transcriptional products. MIR22HG is an oncogenic lncRNA highly expressed in human glioblastoma neurons. Inactivation of MIR22HG via siRNAs has been shown to suppress glioblastoma development in vivo (Han et al., 2020). Most recently, a new branch of regulatory ncRNAs, called circular RNAs (cirRNAs) has been discovered in regulating gene expression. These are single-stranded RNAs covalently looped at their 3’ and 5’ ends with unclear function but have strong associations with numerous diseases.
ncRNAs have great therapeutic potential due to their diverse functionality. There is growing interest in modulating ncRNAs to treat various disorders. ncRNAs are a new and exciting field of research that will revolutionize our understanding of human diseases. No longer considered the “dark matter” or “junk” of the human genome, ncRNAs have expanded the central dogma of molecular biology and are changing the way we view genetic expression.
About the Author
Rachel Chau is a junior at Harvard College concentrating in Neuroscience.
References
Ameres, S. L., & Zamore, P. D. (2013). Diversifying microrna sequence and function. Nature Reviews Molecular Cell Biology, 14(8), 475–488. https://doi.org/10.1038/nrm3611
Baptista, B., Riscado, M., Queiroz, J. A., Pichon, C., & Sousa, F. (2021). Non-coding RNAS: Emerging from the discovery to therapeutic applications. Biochemical Pharmacology, 189, 114469. https://doi.org/10.1016/j.bcp.2021.114469
Djebali, S., Davis, C. A., Merkel, A., Dobin, A., Lassmann, T., Mortazavi, A., Tanzer, A., Lagarde, J., Lin, W., Schlesinger, F., Xue, C., Marinov, G. K., Khatun, J., Williams, B. A., Zaleski, C., Rozowsky, J., Röder, M., Kokocinski, F., Abdelhamid, R. F., … Gingeras, T. R. (2012). Landscape of transcription in human cells. Nature, 489(7414), 101–108. https://doi.org/10.1038/nature11233
International Human Genome Sequencing Consortium. (2004). Finishing the euchromatic sequence of the human genome. Nature, 431(7011), 931–945. https://doi.org/10.1038/nature03001
Mattick, J. S. (2001). Non‐coding RNAS: The Architects of Eukaryotic Complexity. EMBO Reports, 2(11), 986–991. https://doi.org/10.1093/embo-reports/kve230
Nie, J.-H., Li, T.-X., Zhang, X.-Q., & Liu, J. (2019). Roles of non-coding RNAS in normal human brain development, brain tumor, and neuropsychiatric disorders. Non-Coding RNA, 5(2), 36. https://doi.org/10.3390/ncrna5020036
Sartorelli, V., & Lauberth, S. M. (2020). Enhancer RNAS are an important regulatory layer of the epigenome. Nature Structural & Molecular Biology, 27(6), 521–528. https://doi.org/10.1038/s41594-020-0446-0
Xu, J.-zhong, Zhang, J.-lan, & Zhang, W.-guo. (2018). Antisense RNA: The new favorite in genetic research. Journal of Zhejiang University-SCIENCE B, 19(10), 739–749. https://doi.org/10.1631/jzus.b1700594
Long ncRNAs (lncRNAs) have more than 200 nt and constitute most non-coding transcriptional products. MIR22HG is an oncogenic lncRNA highly expressed in human glioblastoma neurons. Inactivation of MIR22HG via siRNAs has been shown to suppress glioblastoma development in vivo (Han et al., 2020). Most recently, a new branch of regulatory ncRNAs, called circular RNAs (cirRNAs) has been discovered in regulating gene expression. These are single-stranded RNAs covalently looped at their 3’ and 5’ ends with unclear function but have strong associations with numerous diseases.
ncRNAs have great therapeutic potential due to their diverse functionality. There is growing interest in modulating ncRNAs to treat various disorders. ncRNAs are a new and exciting field of research that will revolutionize our understanding of human diseases. No longer considered the “dark matter” or “junk” of the human genome, ncRNAs have expanded the central dogma of molecular biology and are changing the way we view genetic expression.
About the Author
Rachel Chau is a junior at Harvard College concentrating in Neuroscience.
References
Ameres, S. L., & Zamore, P. D. (2013). Diversifying microrna sequence and function. Nature Reviews Molecular Cell Biology, 14(8), 475–488. https://doi.org/10.1038/nrm3611
Baptista, B., Riscado, M., Queiroz, J. A., Pichon, C., & Sousa, F. (2021). Non-coding RNAS: Emerging from the discovery to therapeutic applications. Biochemical Pharmacology, 189, 114469. https://doi.org/10.1016/j.bcp.2021.114469
Djebali, S., Davis, C. A., Merkel, A., Dobin, A., Lassmann, T., Mortazavi, A., Tanzer, A., Lagarde, J., Lin, W., Schlesinger, F., Xue, C., Marinov, G. K., Khatun, J., Williams, B. A., Zaleski, C., Rozowsky, J., Röder, M., Kokocinski, F., Abdelhamid, R. F., … Gingeras, T. R. (2012). Landscape of transcription in human cells. Nature, 489(7414), 101–108. https://doi.org/10.1038/nature11233
International Human Genome Sequencing Consortium. (2004). Finishing the euchromatic sequence of the human genome. Nature, 431(7011), 931–945. https://doi.org/10.1038/nature03001
Mattick, J. S. (2001). Non‐coding RNAS: The Architects of Eukaryotic Complexity. EMBO Reports, 2(11), 986–991. https://doi.org/10.1093/embo-reports/kve230
Nie, J.-H., Li, T.-X., Zhang, X.-Q., & Liu, J. (2019). Roles of non-coding RNAS in normal human brain development, brain tumor, and neuropsychiatric disorders. Non-Coding RNA, 5(2), 36. https://doi.org/10.3390/ncrna5020036
Sartorelli, V., & Lauberth, S. M. (2020). Enhancer RNAS are an important regulatory layer of the epigenome. Nature Structural & Molecular Biology, 27(6), 521–528. https://doi.org/10.1038/s41594-020-0446-0
Xu, J.-zhong, Zhang, J.-lan, & Zhang, W.-guo. (2018). Antisense RNA: The new favorite in genetic research. Journal of Zhejiang University-SCIENCE B, 19(10), 739–749. https://doi.org/10.1631/jzus.b1700594
